


畜牧兽医学报 ›› 2025, Vol. 56 ›› Issue (9): 4241-4252.doi: 10.11843/j.issn.0366-6964.2025.09.010
黎梦帆1( ), 李青阳1(), 宋艳雯1(), 宋振辉1,2, 张兴翠1,*()
), 李青阳1(), 宋艳雯1(), 宋振辉1,2, 张兴翠1,*()
收稿日期:2024-11-01
出版日期:2025-09-23
发布日期:2025-09-30
通讯作者:
张兴翠
E-mail:1285530053@qq.com;1852112998@qq.com;962060187@qq.com;zhangxc923@163.com
作者简介:黎梦帆(2003-),女,四川达州人,本科生,主要从事动物病原微生物与免疫研究,E-mail:1285530053@qq.com黎梦帆、李青阳和宋艳雯为同等贡献作者
基金资助:
LI Mengfan1(), LI Qingyang1(), SONG Yanwen1(), SONG Zhenhui1,2, ZHANG Xingcui1,*()
Received:2024-11-01
Online:2025-09-23
Published:2025-09-30
Contact:
ZHANG Xingcui
E-mail:1285530053@qq.com;1852112998@qq.com;962060187@qq.com;zhangxc923@163.com
摘要:
冠状病毒(coronavirus,CoV)是具有包膜的单股正链RNA病毒,作为人和动物的重要致病原,冠状病毒感染主要导致宿主呼吸系统、肝脏、胃肠道以及神经系统出现急性或慢性症状。冠状病毒可感染人类和多种动物,严重危害人体健康和畜牧业发展。绝大多数冠状病毒的刺突(spike,S)蛋白由S1亚基和S2亚基组成,在病毒感染细胞过程中分别负责受体结合和与细胞膜融合,其在病毒的组织或宿主嗜性和毒力等方面发挥着重要作用,是冠状病毒感染细胞的关键蛋白。本文重点对冠状病毒S蛋白的结构和功能进行综述,以期为冠状病毒的入侵机制研究提供参考。
中图分类号:
黎梦帆, 李青阳, 宋艳雯, 宋振辉, 张兴翠. 冠状病毒S蛋白的结构和功能研究进展[J]. 畜牧兽医学报, 2025, 56(9): 4241-4252.
LI Mengfan, LI Qingyang, SONG Yanwen, SONG Zhenhui, ZHANG Xingcui. Structure and Function of Coronavirus S Proteins[J]. Acta Veterinaria et Zootechnica Sinica, 2025, 56(9): 4241-4252.
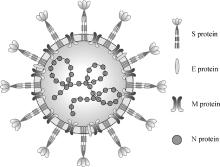
图 1
冠状病毒粒子示意图(彩图在OSID相关材料中提供)"
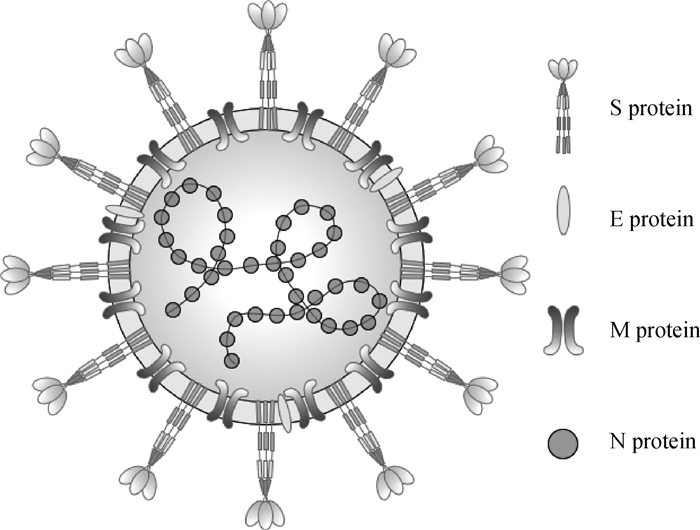

图 2
S蛋白融合前结构和结构域示意图(彩图在OSID相关材料中提供) A. S蛋白结构;B. S蛋白结构域示意图"
表 1
冠状病毒功能性受体以及辅助受体因子"
| 属 Genera | 病毒 Viruses | 宿主 Hosts | 功能性受体 Functional receptors | 辅助性受体 Helper receptors | 参考文献 References |
| 甲型冠状病毒属 Alphacoronaviruses | HCoV-229E | Human | APN | [ | |
| HCoV-NL63 | Porcine | ACE2 | HS | [ | |
| TGEV | APN | SA | [ | ||
| PEDV | APN | SA | [ | ||
| SADS-CoV | Feline | HS、SA | [ | ||
| FECV | APN | SA | [ | ||
| FIPV Ⅰ | [ | ||||
| FIPV Ⅱ | Canine | APN | |||
| CCoV Ⅰ | [ | ||||
| CCoV Ⅱ | APN | ||||
| 乙型冠状病毒属 Betacoronavirus | SARS-CoV | Human | ACE2 | [ | |
| SARS-CoV2 | ACE2 | [ | |||
| MERS-CoV | DPP4 | SA | [ | ||
| HCoV-HKU1 | TMPRSS2 | SA | [ | ||
| HCoV-OC43 | SA | [ | |||
| BCoV | Bovine | APN | [ | ||
| MHV | Murine | CEACAM1 | [ | ||
| 丙型冠状病毒属 Gammacoronavirus | IBV | Avian | SA | [ | |
| 丁型冠状病毒属 Deltacoronavirus | PDCoV | Porcine | APN | [ |

图 3
冠状病毒S原聚体CTD关闭和打开的结构示意图[2](彩图在OSID相关材料中提供) A. 关闭构象;B. 打开构象"
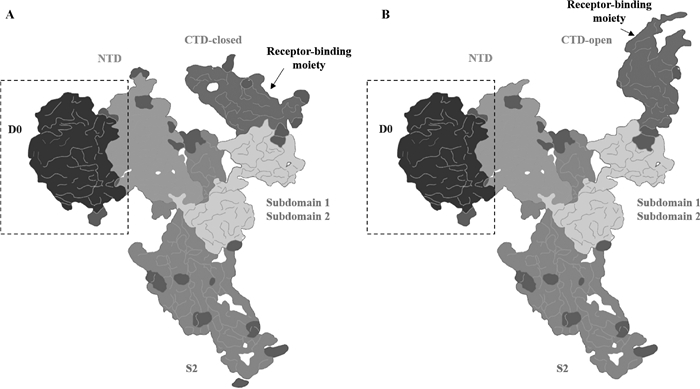
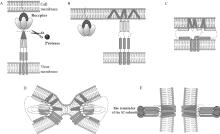
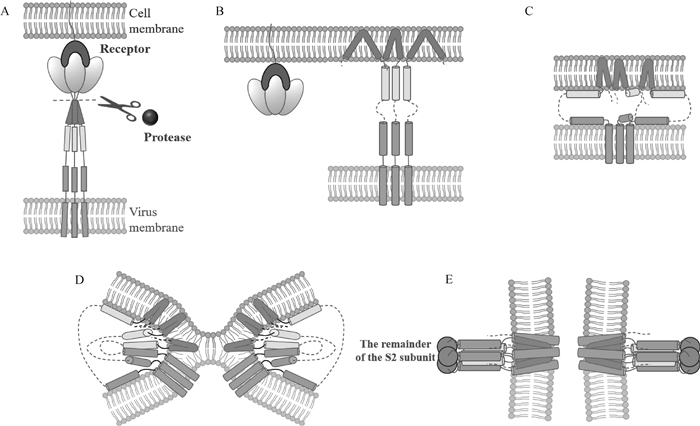
图 4
冠状病毒S蛋白促进病毒进入细胞的膜融合模型[48](彩图在OSID相关材料中提供) S1亚基与受体结合,S蛋白被蛋白酶裂解(图A),S2亚基上的融合肽(FP)暴露并插入宿主细胞膜(图B),七肽重复序列1(heptad repeat,HR1)和HR2折叠形成六螺旋束结构(图C~E),使FP和单通道跨膜锚(TM)对齐,进一步促使病毒膜和细胞膜发生融合(图D、E)"
| 1 |
VAKULENKO Y , DEVIATKIN A , DREXLER J F , et al. Modular evolution of coronavirus genomes[J]. Viruses, 2021, 13 (7): 1270.
doi: 10.3390/v13071270 |
| 2 |
HUANG C Y , DRACZKOWSKI P , WANG Y S , et al. In situ structure and dynamics of an alphacoronavirus spike protein by cryo-ET and cryo-EM[J]. Nat Commun, 2022, 13 (1): 4877.
doi: 10.1038/s41467-022-32588-3 |
| 3 |
SHANG J , ZHENG Y , YANG Y , et al. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins[J]. PLoS Pathog, 2018, 14 (4): e1007009.
doi: 10.1371/journal.ppat.1007009 |
| 4 | LI Z , TOMLINSON A C , WONG A H , et al. The human coronavirus HCoV-229E S-protein structure and receptor binding[J]. Elife, 2019, 8, e1007009. |
| 5 |
WALLS A C , XIONG X , PARK Y J , et al. Unexpected receptor functional mimicry elucidates activation of coronavirus fusion[J]. Cell, 2019, 176 (5): 1026- 1039.e15.
doi: 10.1016/j.cell.2018.12.028 |
| 6 |
CHEN Y , ZHANG Z , LI J , et al. Porcine epidemic diarrhea virus S1 protein is the critical inducer of apoptosis[J]. Virol J, 2018, 15 (1): 170.
doi: 10.1186/s12985-018-1078-4 |
| 7 |
LI F . Receptor recognition mechanisms of coronaviruses: a decade of structural studies[J]. J Virol, 2015, 89 (4): 1954- 1964.
doi: 10.1128/JVI.02615-14 |
| 8 | WRAPP D , MCLELLAN J S . The 3.1-angstrom cryo-electron microscopy structure of the porcine epidemic diarrhea virus spike protein in the prefusion conformation[J]. J Virol, 2019, 93 (23): e00923- 19. |
| 9 |
YU J , QIAO S , GUO R , et al. Cryo-EM structures of HKU2 and SADS-CoV spike glycoproteins provide insights into coronavirus evolution[J]. Nat Commun, 2020, 11 (1): 3070.
doi: 10.1038/s41467-020-16876-4 |
| 10 | GUAN H , WANG Y , PER AČG ULIJA V , et al. Cryo-electron microscopy structure of the swine acute diarrhea syndrome coronavirus spike glycoprotein provides insights into evolution of unique coronavirus spike proteins[J]. J Virol, 2020, 94 (22): e01301- 20. |
| 11 |
WALLS A C , TORTORICI M A , BOSCH B J , et al. Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer[J]. Nature, 2016, 531 (7592): 114- 117.
doi: 10.1038/nature16988 |
| 12 |
WALLS A C , TORTORICI M A , FRENZ B , et al. Glycan shield and epitope masking of a coronavirus spike protein observed by cryo-electron microscopy[J]. Nat Struct Mol Biol, 2016, 23 (10): 899- 905.
doi: 10.1038/nsmb.3293 |
| 13 |
YANG T J , CHANG Y C , KO T P , et al. Cryo-EM analysis of a feline coronavirus spike protein reveals a unique structure and camouflaging glycans[J]. Proc Natl Acad Sci U S A, 2020, 117 (3): 1438- 1446.
doi: 10.1073/pnas.1908898117 |
| 14 |
ZHANG K , LI S , PINTILIE G , et al. A 3.4-Å cryo-electron microscopy structure of the human coronavirus spike trimer computationally derived from vitrified NL63 virus particles[J]. QRB Discov, 2020, 1, e11.
doi: 10.1017/qrd.2020.16 |
| 15 |
LI Y , WANG T , ZHANG J , et al. Exploring the regulatory function of the N-terminal domain of SARS-CoV-2 spike protein through molecular dynamics simulation[J]. Adv Theory Simul, 2021, 4 (10): 2100152.
doi: 10.1002/adts.202100152 |
| 16 |
SHANG J , WAN Y , LIU C , et al. Structure of mouse coronavirus spike protein complexed with receptor reveals mechanism for viral entry[J]. PLoS Pathog, 2020, 16 (3): e1008392.
doi: 10.1371/journal.ppat.1008392 |
| 17 |
LING A J W , CHANG L S , BABJI A S , et al. Review of sialic acid's biochemistry, sources, extraction and functions with special reference to edible bird's nest[J]. Food Chem, 2022, 367, 130755.
doi: 10.1016/j.foodchem.2021.130755 |
| 18 |
DIEP N V , NORIMINE J , SUEYOSHI M , et al. Novel porcine epidemic diarrhea virus (PEDV) variants with large deletions in the spike (S) gene coexist with PEDV strains possessing an intact S gene in domestic pigs in Japan: A new disease situation[J]. PLoS One, 2017, 12 (1): e0170126.
doi: 10.1371/journal.pone.0170126 |
| 19 |
PENG G , XU L , LIN Y L , et al. Crystal structure of bovine coronavirus spike protein lectin domain[J]. J Biol Chem, 2012, 287 (50): 41931- 41938.
doi: 10.1074/jbc.M112.418210 |
| 20 |
YOU R , LIU K , HUANG M , et al. Identification and comparison of the sialic acid-binding domain characteristics of avian coronavirus infectious bronchitis virus spike protein[J]. J Virol, 2023, 97 (5): e0048923.
doi: 10.1128/jvi.00489-23 |
| 21 |
QIAO M , LIN L , XIA K , et al. Recent advances in biotechnology for heparin and heparan sulfate analysis[J]. Talanta, 2020, 219, 121270.
doi: 10.1016/j.talanta.2020.121270 |
| 22 | NASKALSKA A , DABROWSKA A , SZCZEPANSKI A , et al. Membrane protein of human coronavirus NL63 is responsible for interaction with the adhesion receptor[J]. J Virol, 2019, 93 (19): e00355- 19. |
| 23 |
YANG Y L , WANG B , LI W , et al. Functional dissection of the spike glycoprotein S1 subunit and identification of cellular cofactors for regulation of swine acute diarrhea syndrome coronavirus entry[J]. J Virol, 2024, 98 (4): e0013924.
doi: 10.1128/jvi.00139-24 |
| 24 | DESMARETS L M B , THEUNS S , ROUKAERTS I D M , et al. Role of sialic acids in feline enteric coronavirus infections[J]. J Gen Virol, 2014, 95 (Pt 9): 1911- 1918. |
| 25 |
COOK S , CASTILLO D , WILLIAMS S , et al. Serotype Ⅰ and Ⅱ feline coronavirus replication and gene expression patterns of feline cells-building a better understanding of serotype I FIPV biology[J]. Viruses, 2022, 14 (7): 1356.
doi: 10.3390/v14071356 |
| 26 |
CERRACCHIO C , SERRA F , AMOROSO M G , et al. Canine coronavirus activates Aryl hydrocarbon receptor during in vitro infection[J]. Viruses, 2022, 14 (11): 2437.
doi: 10.3390/v14112437 |
| 27 | GUO H , HU B J , YANG X L , et al. Evolutionary arms race between virus and host drives genetic diversity in bat severe acute respiratory syndrome-related coronavirus spike genes[J]. J Virol, 2020, 94 (20): e00902- 20. |
| 28 |
LAN J , GE J , YU J , et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor[J]. Nature, 2020, 581 (7807): 215- 220.
doi: 10.1038/s41586-020-2180-5 |
| 29 |
ALAOFI A L . Exploring structural dynamics of the MERS-CoV receptor DPP4 and mutant DPP4 receptors[J]. J Biomol Struct Dyn, 2022, 40 (2): 752- 763.
doi: 10.1080/07391102.2020.1818626 |
| 30 |
SAUNDERS N , FERNANDEZ I , PLANCHAIS C , et al. TMPRSS2 is a functional receptor for human coronavirus HKU1[J]. Nature, 2023, 624 (7990): 207- 214.
doi: 10.1038/s41586-023-06761-7 |
| 31 |
TANG G , LIU Z , CHEN D . Human coronaviruses: Origin, host and receptor[J]. J Clin Virol, 2022, 155, 105246.
doi: 10.1016/j.jcv.2022.105246 |
| 32 |
JI W , PENG Q , FANG X , et al. Structures of a deltacoronavirus spike protein bound to porcine and human receptors[J]. Nat Commun, 2022, 13 (1): 1467.
doi: 10.1038/s41467-022-29062-5 |
| 33 |
SHI J , SHI Y , XIU R , et al. Identification of a novel neutralizing epitope on the N-terminal domain of the human coronavirus 229E spike protein[J]. J Virol, 2022, 96 (4): e0195521.
doi: 10.1128/jvi.01955-21 |
| 34 | SHANG J , ZHENG Y , YANG Y , et al. Cryo-electron microscopy structure of porcine deltacoronavirus spike protein in the prefusion state[J]. J Virol, 2018, 92 (4): e01556- 17. |
| 35 |
PAN Y , TIAN X , QIN P , et al. Discovery of a novel swine enteric alphacoronavirus (SeACoV) in southern China[J]. Vet Microbiol, 2017, 211, 15- 21.
doi: 10.1016/j.vetmic.2017.09.020 |
| 36 |
LI F . Structure, function, and evolution of coronavirus spike proteins[J]. Annu Rev Virol, 2016, 3 (1): 237- 261.
doi: 10.1146/annurev-virology-110615-042301 |
| 37 |
LI C , SU M , YIN B , et al. Integrin αvβ3 enhances replication of porcine epidemic diarrhea virus on Vero E6 and porcine intestinal epithelial cells[J]. Vet Microbiol, 2019, 237, 108400.
doi: 10.1016/j.vetmic.2019.108400 |
| 38 |
WARDEH M , BAYLIS M , BLAGROVE M S C . Predicting mammalian hosts in which novel coronaviruses can be generated[J]. Nat Commun, 2021, 12 (1): 780.
doi: 10.1038/s41467-021-21034-5 |
| 39 |
WANG S , XU C , SHI J , et al. Regulatory effect and mechanism of APN gene on porcine epidemic diarrhea virus resistance[J]. Gene, 2021, 775, 145448.
doi: 10.1016/j.gene.2021.145448 |
| 40 |
PIZZANELLI S , FORTE C , PINZINO C , et al. Copper(Ⅱ) complexes with peptides based on the second cell binding site of fibronectin: metal coordination and ligand exchange kinetics[J]. Phys Chem Chem Phys, 2016, 18 (5): 3982- 3994.
doi: 10.1039/C5CP05798A |
| 41 |
CASTILLO G , MORA-DíAZ J C , NELLI R K , et al. Human air-liquid-interface organotypic airway cultures express significantly more ACE2 receptor protein and are more susceptible to HCoV-NL63 infection than monolayer cultures of primary respiratory epithelial cells[J]. Microbiol Spectr, 2022, 10 (4): e0163922.
doi: 10.1128/spectrum.01639-22 |
| 42 | PECK K M , SCOBEY T , SWANSTROM J , et al. Permissivity of dipeptidyl peptidase 4 orthologs to Middle East respiratory syndrome coronavirus is governed by glycosylation and other complex determinants[J]. J Virol, 2017, 91 (19): e00534- 17. |
| 43 | PALLESEN J , WANG N , CORBETT K S , et al. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen[J]. Proc Natl Acad Sci U S A, 2017, 114 (35): E7348- E7357. |
| 44 |
KIRCHDOERFER R N , WANG N , PALLESEN J , et al. Stabilized coronavirus spikes are resistant to conformational changes induced by receptor recognition or proteolysis[J]. Sci Rep, 2018, 8 (1): 15701.
doi: 10.1038/s41598-018-34171-7 |
| 45 |
GUI M , SONG W , ZHOU H , et al. Cryo-electron microscopy structures of the SARS-CoV spike glycoprotein reveal a prerequisite conformational state for receptor binding[J]. Cell Res, 2017, 27 (1): 119- 129.
doi: 10.1038/cr.2016.152 |
| 46 |
WALLS A C , TORTORICI M A , SNIJDER J , et al. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion[J]. Proc Natl Acad Sci U S A, 2017, 114 (42): 11157- 11162.
doi: 10.1073/pnas.1708727114 |
| 47 |
SONG X , SHI Y , DING W , et al. Cryo-EM analysis of the HCoV-229E spike glycoprotein reveals dynamic prefusion conformational changes[J]. Nat Commun, 2021, 12 (1): 141.
doi: 10.1038/s41467-020-20401-y |
| 48 |
ALIPER E T , EFREMOV R G . Inconspicuous yet indispensable: The coronavirus spike transmembrane domain[J]. Int J Mol Sci, 2023, 24 (22): 16421.
doi: 10.3390/ijms242216421 |
| 49 |
KIELIAN M . Mechanisms of virus membrane fusion proteins[J]. Annu Rev Virol, 2014, 1 (1): 171- 189.
doi: 10.1146/annurev-virology-031413-085521 |
| 50 |
SHI W , CAI Y , ZHU H , et al. Cryo-EM structure of SARS-CoV-2 postfusion spike in membrane[J]. Nature, 2023, 619 (7969): 403- 409.
doi: 10.1038/s41586-023-06273-4 |
| 51 |
BASSO L G M , ZERAIK A E , FELIZATTI A P , et al. Membranotropic and biological activities of the membrane fusion peptides from SARS-CoV spike glycoprotein: The importance of the complete internal fusion peptide domain[J]. Biochim Biophys Acta Biomembr, 2021, 1863 (11): 183697.
doi: 10.1016/j.bbamem.2021.183697 |
| 52 |
WALLS A C , PARK Y J , TORTORICI M A , et al. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein[J]. Cell, 2020, 181 (2): 281- 92.e6.
doi: 10.1016/j.cell.2020.02.058 |
| 53 |
MAEDA D , TIAN D , YU H , et al. Killed whole-genome reduced-bacteria surface-expressed coronavirus fusion peptide vaccines protect against disease in a porcine model[J]. Proc Natl Acad Sci U S A, 2021, 118 (18): e2025622118.
doi: 10.1073/pnas.2025622118 |
| 54 |
DACON C , TUCKER C , PENG L , et al. Broadly neutralizing antibodies target the coronavirus fusion peptide[J]. Science, 2022, 377 (6607): 728- 735.
doi: 10.1126/science.abq3773 |
| 55 |
LOW J S , JERAK J , TORTORICI M A , et al. ACE2-binding exposes the SARS-CoV-2 fusion peptide to broadly neutralizing coronavirus antibodies[J]. Science, 2022, 377 (6607): 735- 742.
doi: 10.1126/science.abq2679 |
| 56 |
SUN X , YI C , ZHU Y , et al. Neutralization mechanism of a human antibody with pan-coronavirus reactivity including SARS-CoV-2[J]. Nat Microbiol, 2022, 7 (7): 1063- 1074.
doi: 10.1038/s41564-022-01155-3 |
| 57 |
MILLET J K , WHITTAKER G R . Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis[J]. Virus Res, 2015, 202, 120- 34.
doi: 10.1016/j.virusres.2014.11.021 |
| 58 | YAN L , MENG B , XIANG J , et al. Crystal structure of the post-fusion core of the human coronavirus 229E spike protein at 1.86 Å resolution[J]. Acta Crystallogr D Struct Biol, 2018, 74 (Pt 9): 841- 851. |
| 59 |
WANG F , YANG G , YAN L . Crystal Structures of Fusion Cores from CCoV-HuPn-2018 and SADS-CoV[J]. Viruses, 2024, 16 (2): 272.
doi: 10.3390/v16020272 |
| 60 |
WESTERFIELD J M , BARRERA F N . Membrane receptor activation mechanisms and transmembrane peptide tools to elucidate them[J]. J Biol Chem, 2020, 295 (7): 1792- 1814.
doi: 10.1074/jbc.REV119.009457 |
| 61 |
KUMAR P , BHARDWAJ T , GARG N , et al. Microsecond simulations and CD spectroscopy reveals the intrinsically disordered nature of SARS-CoV-2 spike-C-terminal cytoplasmic tail (residues 1242-1273) in isolation[J]. Virology, 2022, 566, 42- 55.
doi: 10.1016/j.virol.2021.11.005 |
| 62 |
SADASIVAN J , SINGH M , SARMA J D . Cytoplasmic tail of coronavirus spike protein has intracellular targeting signals[J]. J Biosci, 2017, 42 (2): 231- 244.
doi: 10.1007/s12038-017-9676-7 |
| 63 |
WU Z , ZHANG Z , WANG X , et al. Palmitoylation of SARS-CoV-2 S protein is essential for viral infectivity[J]. Signal Transduct Target Ther, 2021, 6 (1): 231.
doi: 10.1038/s41392-021-00651-y |
| 64 |
LUO Y , TAN C W , XIE S Z , et al. Identification of ZDHHC17 as a potential drug target for swine acute diarrhea syndrome coronavirus infection[J]. mBio, 2021, 12 (5): e0234221.
doi: 10.1128/mBio.02342-21 |
| 65 |
GELHAUS S , THAA B , ESCHKE K , et al. Palmitoylation of the Alphacoronavirus TGEV spike protein S is essential for incorporation into virus-like particles but dispensable for S-M interaction[J]. Virology, 2014, 464-465, 397- 405.
doi: 10.1016/j.virol.2014.07.035 |
| 66 |
KUMAR P , BHARDWAJ A , MUKHERJEE B , et al. Coronaviruses spike glycoprotein endodomains: The sequence and structure-based comprehensive study[J]. Protein Sci, 2023, 32 (11): e4804.
doi: 10.1002/pro.4804 |
| 67 |
CHENG Y R , LI X , ZHAO X , et al. Cell entry of animal coronaviruses[J]. Viruses, 2021, 13 (10): 1977.
doi: 10.3390/v13101977 |
| 68 |
JOHNSON B A , XIE X , BAILEY A L , et al. Loss of furin cleavage site attenuates SARS-CoV-2 pathogenesis[J]. Nature, 2021, 591 (7849): 293- 299.
doi: 10.1038/s41586-021-03237-4 |
| 69 |
WANG Z , ZHONG K , WANG G , et al. Loss of furin site enhances SARS-CoV-2 spike protein pseudovirus infection[J]. Gene, 2023, 856, 147144.
doi: 10.1016/j.gene.2022.147144 |
| 70 | ZEHR J D , KOSAKOVSKY POND S L , MILLET J K , et al. Natural selection differences detected in key protein domains between non-pathogenic and pathogenic feline coronavirus phenotypes[J]. Virus Evol, 2023, 9 (1): 523607. |
| 71 |
KIM J , YOON J , PARK J E . Furin cleavage is required for swine acute diarrhea syndrome coronavirus spike protein-mediated cell-cell fusion[J]. Emerg Microbes Infect, 2022, 11 (1): 2176- 2183.
doi: 10.1080/22221751.2022.2114850 |
| 72 |
BONNIN A , DANNEELS A , DUBUISSON J , et al. HCoV-229E spike protein fusion activation by trypsin-like serine proteases is mediated by proteolytic processing in the S2' region[J]. J Gen Virol, 2018, 99 (7): 908- 912.
doi: 10.1099/jgv.0.001074 |
| 73 |
LIN F , ZHANG H , LI L , et al. PEDV: Insights and advances into types, function, structure, and receptor recognition[J]. Viruses, 2022, 14 (8): 1744.
doi: 10.3390/v14081744 |
| 74 |
WICHT O , LI W , WILLEMS L , et al. Proteolytic activation of the porcine epidemic diarrhea coronavirus spike fusion protein by trypsin in cell culture[J]. J Virol, 2014, 88 (14): 7952- 7961.
doi: 10.1128/JVI.00297-14 |
| 75 |
SUN M , MA J , YU Z , et al. Identification of two mutation sites in spike and envelope proteins mediating optimal cellular infection of porcine epidemic diarrhea virus from different pathways[J]. Vet Res, 2017, 48 (1): 44.
doi: 10.1186/s13567-017-0449-y |
| [1] | 刘君君, 郭东辉, 刘缓缓, 宋润泽, 赵赛娅, 杨钧尧, 魏战勇, 项玉强, 陈丽颖. 基于免疫磁珠的智能手机辅助比色传感平台用于PDCoV/TGEV IgG抗体的快速检测[J]. 畜牧兽医学报, 2025, 56(9): 4559-4571. |
| [2] | 余秋蓉, 蔡旭航, 何艺, 李基棕, 毛立, 许信刚, 李彬. 一株羊冠状病毒的分离鉴定及全基因组序列分析[J]. 畜牧兽医学报, 2025, 56(9): 4604-4614. |
| [3] | 马思琪, 吕雯雯, 陈俊贞, 李建林, 刘昱成, 关团, 丁剑, 刘浩然, 叶鸿艳, 杨莉, 付强, 史慧君. 牛肠病毒VP1基因重组腺病毒的构建及其对小鼠的免疫原性评价[J]. 畜牧兽医学报, 2025, 56(9): 4615-4625. |
| [4] | 常铄, 孙秀柱, 任战军, 王淑辉. 家兔基因组学研究进展[J]. 畜牧兽医学报, 2025, 56(8): 3578-3590. |
| [5] | 陶丽寒, 林翠, 吴诚诚, 康昭风, 黄建珍. 猪丁型冠状病毒编码蛋白结构与功能研究进展[J]. 畜牧兽医学报, 2025, 56(8): 3678-3689. |
| [6] | 刘莎, 杨彩春, 张晓雨, 陈琼, 刘雄, 陈洪波, 周焕焕, 史良玉. 基于80K SNP芯片的梅花星猪群体遗传结构解析及全基因组连续纯合片段特征研究[J]. 畜牧兽医学报, 2025, 56(8): 3749-3760. |
| [7] | 任千姿, 张佰忠, 王真勍, 王向林, 龚颖, 胡仁科, 浦亚斌, 苏鹏, 李业芳, 马月辉, 李昊帮, 蒋琳. 基于全基因组重测序对武雪山羊的遗传进化分析[J]. 畜牧兽医学报, 2025, 56(8): 3787-3801. |
| [8] | 李茜, 高欢, 符爽, 锁卓, 代悦, 陈晨, 李荣天, 冷静. 消化道厌氧真菌及与其他微生物的互作关系[J]. 畜牧兽医学报, 2025, 56(7): 3096-3106. |
| [9] | 向令娴, 季倩宇, 单新新, 李琳. 细菌双组分系统的耐药性及致病性研究进展[J]. 畜牧兽医学报, 2025, 56(7): 3116-3128. |
| [10] | 张嘉良, 黄畅, 杨永林, 杨华, 白文林, 马月辉, 赵倩君. 基于50K液相芯片的中国绵羊群体遗传结构与羊毛性状选择信号分析[J]. 畜牧兽医学报, 2025, 56(7): 3164-3176. |
| [11] | 潘华, 孙磊, 张军, 刘莉莉, 马文刚, 曹爱智, 吕明斌. 胆汁酸抑菌机制及结构改性的研究进展[J]. 畜牧兽医学报, 2025, 56(6): 2546-2554. |
| [12] | 董娇娇, 丁虹, 张寅梁, 张冉, 刘华格, 臧素敏, 张振红, 周荣艳, 李兰会. 鸡白痢沙门菌感染太行鸡盲肠菌群的差异及功能分析[J]. 畜牧兽医学报, 2025, 56(6): 2741-2751. |
| [13] | 王勤倩, 高振东, 陆颖, 马若珊, 邓卫东, 和晓明. 全基因组重测序在中国地方黄牛上的研究进展[J]. 畜牧兽医学报, 2025, 56(5): 2026-2037. |
| [14] | 秦小霞, 甘海清, 佘高进, 刘勇, 黄兴国, 陈丽蓉, 杨玲媛. 油茶籽粕的活性成分、生物学功能及其在畜禽生产中的应用研究进展[J]. 畜牧兽医学报, 2025, 56(5): 2070-2081. |
| [15] | 吴芊卉, 张愉, 张桃妮, 磨美兰. 脂筏参与冠状病毒感染的机制及其应用的研究进展[J]. 畜牧兽医学报, 2025, 56(5): 2112-2122. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||